Research Progress and Preclinical Prospects of Plant-Derived Extracellular Vesicles in Targeted Delivery of Antitumor Drugs

Introduction
Extracellular vesicles (EVs) have emerged as pivotal mediators of intercellular communication, facilitating the transfer of bioactive molecules such as proteins, lipids, nucleic acids, and metabolites between cells. This unique capacity enables EVs to regulate diverse physiological and pathological processes, including tumor progression and immune modulation.1,2 In cancer, EVs are not only involved in the crosstalk among tumor cells and their microenvironment but also hold promise as natural carriers for targeted drug delivery, owing to their inherent biocompatibility, low immunogenicity, and cerapeutics which often suffer from poor targeting and significant systemic toxicity.
Among the diverse sources of EVs, plant-derived extracellular vesicles (PDEVs) have garnered increasing attention due to their abundant availability, safety profile, and favorable biological properties. Unlike mammalian cell-derived EVs, PDEVs can be isolated from various plant tissues such as fruit juice, roots, and leaves, providing a costeffective and scalable source.1 PDEVs are primarily isolated via two distinct approaches: (1) mechanical disruption of plant tissues-employing techniques such as tissue squeezing, sonication, or high-pressure homogenization-to liberate intracellular vesicles; and (2) isolation of naturally secreted vesicles through gentle, non-disruptive extraction methods-including differential ultracentrifugation and affinity-based purification-to preserve their native structural integrity and functional bioactivity.3 Importantly, PDEVs exhibit low immunogenicity and excellent biocompatibility, minimizing adverse immune reactions upon administration. Studies have demonstrated that PDEVs can be engineered or loaded with anticancer agents to achieve targeted delivery, enhancing therapeutic efficacy while reducing off-target effects.4,5 For instance, Isodon rubescens-derived vesicles modified to deliver doxorubicin showed promising oraltargeted therapeutic effects in colitis-associated cancer models, highlighting the potential of PDEVs as drug delivery vectors in oncology.5
The tumor microenvironment (TME) is a dynamic and highly heterogeneous ecosystem that critically influences tumorigenesis, progression, immune evasion, and the acquisition of therapeutic resistance. Comprising not only malignant cells but also diverse stromal and immune cell populations, extracellular matrix components, and soluble factors, the TME is characterized by distinctive biochemical features-including hypoxia, extracellular acidosis, and elevated glutathione concentrations.6,7 Traditional chemotherapy often fails to effectively target this intricate ecosystem, resulting in suboptimal drug accumulation and severe systemic toxicity.8,9 Moreover, the immunosuppressive nature of the TME facilitates tumor escape from immune surveillance, further complicating treatment.10,11 Therefore, the development of innovative, TME-responsive drug delivery systems is imperative to enhance tumor specificity, improve drug penetration, and modulate the immunological landscape to potentiate antitumor responses.12,13 In this context, PDEVs offer a promising solution to the challenges posed by the TME and conventional chemotherapy. Their natural origin and intrinsic bioactivities allow them to interact with and potentially remodel the TME, enhancing drug delivery and immune activation.1,4 Furthermore, the ability to modify PDEVs to carry chemotherapeutic agents or nucleic acids enables the design of multifunctional nanoplatforms that can respond to specific TME stimuli such as pH, redox potential, or enzymatic activity, thereby achieving controlled and targeted drug release.5,8 The low immunogenicity and biocompatibility of PDEVs also support repeated administrations, which is essential for chronic cancer therapy regimens. Collectively, these attributes underscore the potential of PDEVs as ideal carriers for precision antitumor drug delivery, capable of overcoming the limitations of existing therapies.
Given the rapid advances in nanomedicine and the accumulating evidence supporting the therapeutic potential of PDEVs, a comprehensive synthesis of current research is warranted. This review aims to systematically summarize the progress in the application of plant-derived extracellular vesicles for targeted delivery of antitumor drugs, elucidate their mechanisms of action within the tumor microenvironment, and discuss their preclinical prospects and challenges. By integrating insights from recent in vitro and in vivo studies, including investigations into the bioactive compounds of plants such as Momordica charantia and Isodon rubescens, as well as advances in EV engineering and tumor microenvironment modulation, we seek to provide a critical perspective on the translational potential of PDEVs in cancer therapy.1,4,5 Ultimately, this review endeavors to foster the development of novel PDEVs-based strategies that contribute to more effective, safe, and personalized antitumor treatments.
Biological Characteristics and Extraction Techniques of PDEVs
Composition and Structural Characteristics of PDEVs
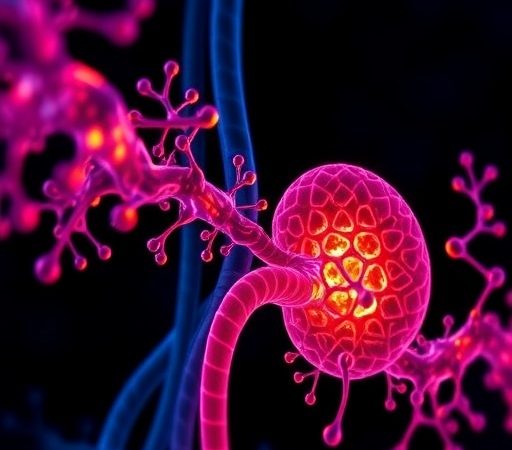
Plant cells have been observed to secrete vesicles that are biogenically and morphologically comparable to mammalian-derived EVs.14 Plant exosome-like nanovesicles, plant-derived nanovesicles and edible plant-derived nanovesicles are the common terms used in different literature to describe nano and micro-sized vesicles (50–1000 nm) isolated from plants. Throughout this review, we will refer to these vesicles as PDEVs. Although the biogenesis of PDEVs is not well-studied, evidence showed that there are at least three different pathways15,16(Figure 1A). (1) The multivesicular body (MVB) pathway is widely considered the main biogenetic route for PDEVs.17–20 As early as 1967, Halperin et al described the formation of vesicles from carrots through this pathway. In this process, MVBs merge with the plasma membrane, leading to the exocytotic release of intraluminal vesicles, known as microvesicles (MVs), into the extracellular environment.17,21 Later research confirmed that extracellular vesicles from Arabidopsis thaliana are also produced and secreted via the MVB pathway.22 (2) The exocyst-positive organelle (EXPO) pathway involves a unique, spherical, double-membrane organelle found only in plant cells. When EXPO fuses with the plasma membrane, it directly secretes microvesicles into the extracellular space.23 This pathway is mainly activated under abiotic or biotic stress conditions and has been experimentally demonstrated in both Arabidopsis and Nicotiana tabacum.24,25 (3) The vacuolar pathway differs from the regulated secretion mechanisms of the MVB and EXPO pathways, representing a context-dependent, stress-induced method of PDEV formation. During pathogen attacks, vacuolar hydrolases and antimicrobial agents are directed to the plasma membrane, promoting the release of defense-related vesicular contents outside the cell.24 Although the central vacuole forms through the fusion of MVB-derived prevacuolar compartments,26 its role in extracellular vesicle release is considered a secondary, inducible process rather than a constant biogenetic pathway. PDEVs are nanoscale vesicles secreted by plant cells, characterized by a lipid bilayer membrane that encapsulates a complex cargo of biomolecules, including lipids, proteins, nucleic acids, and diverse plant metabolites (Figure 1B). This intricate composition endows PDEVs with inherent biological activities and remarkable stability, making them promising candidates for therapeutic applications, especially in targeted drug delivery. Lipid profiling of PDEVs reveals a predominance of fatty acids such as palmitic acid, linoleic acid, and cis-vaccenic acid, which contribute to the vesicles’ membrane integrity and fluidity, thereby ensuring the protection and controlled release of encapsulated agents.27 Proteomic analyses demonstrate that PDEVs harbor proteins associated with extracellular structures and functions, reflecting their role in intercellular communication and plant physiological processes; for example, vesicles derived from Stevia rebaudiana are enriched in extracellular matrix-related proteins, whereas those from Vaccaria hispanica contain more cytosolic proteins, indicating source-dependent compositional variability.27 Additionally, nucleic acids, including small RNAs such as microRNAs and siRNAs, are integral components of PDEVs, enabling them to modulate gene expression in recipient cells and mediate cross-kingdom communication.28 Beyond these biomolecules, PDEVs carry a variety of secondary metabolites unique to their botanical origins, such as polyphenols and other bioactive compounds, which contribute to their pharmacological properties including anti-inflammatory, antioxidant, and antitumor effects.29,30
 |
Figure 1 The biological origin and functional structure of PDEVs. (A) Three pathways of the biological origin of PDEVs. (1) PDEVs associated with tetraspanin 8/tetraspanin 9 (TET8/TET9) are secreted from multivesicular bodies (MVBs). (2) Another subpopulation of PDEVs is derived from exocyst-positive organelles (EXPO). (3) There might be other subpopulations of PDEVs derived from the vacuoles and/or autophagosomes. (B) Schematic representation of PDEVs contents and their relation to PDEVs functions. The composition of PDEVs include lipids, proteins, nucleic acids and secondary metabolites. |
Structurally, PDEVs exhibit a lipid bilayer membrane architecture closely resembling that of animal-derived extracellular vesicles, with a characteristic spherical or cup-shaped morphology observed under electron microscopy.27,31 This membrane structure is crucial for maintaining vesicle stability in diverse physiological environments, protecting the cargo from enzymatic degradation, and facilitating targeted delivery to specific cells or tissues. The bilayer composition not only ensures vesicle integrity but also allows for surface modifications that can enhance targeting specificity and cellular uptake.32,33 The similarity in membrane composition between plant and animal EVs supports the feasibility of PDEVs as biocompatible nanocarriers with low immunogenicity and high permeability, capable of crossing biological barriers such as the intestinal epithelium and even the blood-brain barrier.32 Moreover, the membrane’s lipid constituents contribute to the vesicles’ natural targeting capability, enabling selective interaction with recipient cells and improving drug delivery efficiency.33 The presence of various membrane proteins and glycoproteins further facilitates vesicle recognition and uptake via receptor-mediated endocytosis or fusion, underscoring the sophisticated biological functionality of PDEVs.31
The stability of PDEVs is a critical determinant for their research reproducibility and translational potential. Currently, no standardized guidelines exist for PDEV storage, and reported conditions vary considerably across studies. Short-term storage at 4°C maintains PDEV integrity for 3–14 days, with gradual aggregation reported beyond 7 days for certain sources such as ginger and bitter melon.34 −20°C storage is generally discouraged due to freeze-thaw-induced damage, although single-use aliquots may preserve stability for up to 1 month.35 −80°C is the most widely adopted condition for long-term preservation, with multiple studies demonstrating stability for 3–6 months in PBS without significant changes in size, concentration, or biological activity.36 Lyophilization combined with cryoprotectants such as trehalose or sucrose offers a promising strategy for ambient-temperature storage, though formulation optimization remains an active area of investigation. Notably, the majority of stability studies focus on physicochemical parameters, with limited data on the preservation of functional activity over extended periods. Future efforts should prioritize the establishment of standardized stability assessment protocols, the development of lyophilized formulations, and the generation of long-term stability data exceeding 12 months to support clinical translation.
In summary, the composition of PDEVs is a complex amalgam of lipids, proteins, nucleic acids, and plant-specific metabolites, all encapsulated within a stable lipid bilayer membrane structurally analogous to animal EVs. This unique combination imparts PDEVs with intrinsic biological activities, stability, and targeting abilities, making them highly suitable as natural nanocarriers for the delivery of antitumor drugs and other therapeutic agents. Understanding these compositional and structural characteristics is fundamental for advancing the development of PDEVs-based drug delivery systems and optimizing their clinical translation.30,31,33
Isolation of PDVs
Before the formal isolation process, several physical techniques can be utilized to disrupt plant tissues-referred to as soft tissue disruption-and to extract juice from various plant components, including fruits and stems. Such techniques encompass grinding, squeezing, and blending.37,38 In addition to obtaining juice from plant tissues rich in liquid content, apoplast washing fluid can be extracted through the method of vacuum infiltration.39,40 The apoplast refers to the extracellular region outside the plasma membrane, encompassing both the intercellular spaces and the cell wall. Initially, an infiltration fluid is introduced into the plant tissue-commonly leaves or roots-by creating a pressure differential, often achieved through manual pressure. Subsequently, the infiltrated plant material is subjected to low-speed centrifugation, typically ranging from 700 to 5000 × g, to recover the apoplast washing fluid. Notably, it has been documented that root exudates from hydroponically cultivated plants can be harvested for the isolation of PDVs.41 This process involves substituting the nutrient solution with ultrapure water to facilitate the collection of exudates. Subsequently, the solution containing the exudates is collected for the isolation of PDVs.
The extraction and purification of PDEVs primarily rely on classical techniques such as differential centrifugation, density gradient ultracentrifugation, ultrafiltration size exclusion and precipitation methods. These methods are applied to plant materials including juice, roots, rhizomes, or fruit pulp, aiming to isolate intact vesicles with high purity and functional integrity. Differential centrifugation is widely used as the initial step, involving sequential centrifugation at increasing speeds to remove cell debris and larger particles, followed by ultracentrifugation to pellet the vesicles. The differential centrifugation method is simple to operate and suitable for large-volume samples, but it has a relatively low purity and takes a long time. High shear force may damage the integrity of vesicles.42 To further refine the purity and separate specific EV subclasses, density gradient ultracentrifugation is employed, which separates vesicles based on their buoyant density, allowing for the enrichment of vesicles with particular characteristics. This method has high separation purity and can distinguish different subgroups of vesicles. However, it is cumbersome to operate, takes a long time, has a low recovery rate, and is difficult to be scaled up. Ultrafiltration techniques, often combined with size exclusion chromatography, are also utilized to concentrate and purify vesicles by size exclusion. They can maintain the vesicle structure and activity well, with a high recovery rate; however, the ultrafiltration membrane is prone to clogging, the size exclusion method has a small processing capacity, and it is unable to effectively distinguish similar-sized lipoproteins and other impurities.43 For instance, isolation from apoplastic washing fluid (AWF) of Arabidopsis thaliana leaves involves clean AWF collection followed by differential centrifugation and density gradient steps, with immunoaffinity capture providing the highest specificity when suitable markers and antibodies are available.44 The precipitation method is the simplest to operate, does not require special equipment, has a high throughput, but has the lowest purity. It is prone to co-precipitate proteins, polysaccharides and polymers themselves, which can interfere with downstream analysis.43 Enzymatic pulping approaches, such as the Phyto-EVpure method, have been optimized for medicinal plants like ginger, Morus alba leaves, and Isatis indigotica roots, which improve yield and purity compared to mechanical grinding, and are applicable to fresh, frozen, or dried plant materials.45 The choice of method often depends on the plant source, tissue type, and intended downstream applications. For example, isolation from hairy root cultures of medicinal plants Salvia dominica and S. sclarea uses differential ultracentrifugation combined with size exclusion chromatography, enabling characterization of proteomic and metabolomic cargo relevant to therapeutic potential.46,47 Tobacco-derived vesicles have been isolated using two different methods, revealing differences in vesicle aggregation and uptake efficiency, highlighting the impact of isolation techniques on vesicle properties.48 Furthermore, emerging platforms such as magnetic bead-based depletion of impurities using Concanavalin A-coated beads have demonstrated enhanced purity and functional uptake of liquorice-derived EVs, indicating the importance of removing contaminants like pathogenic bacteria during purification.49
Importantly, the quality and function of PDEVs are significantly influenced by the plant species, tissue type, and even the harvest time. Studies have shown that the maturity stage of the plant or fruit affects the physicochemical and biological properties of isolated EVs. For example, orange-derived EVs from unripe and ripe fruits exhibited superior delivery performance for curcumin compared to those from overripe fruits, suggesting that vesicle composition and cargo loading capacity vary with fruit maturity.50 Similarly, the timing of extraction from Isodon rubescens (a member of the genus Isodon) affects the drug-loading capability of the vesicles, indicating that harvesting conditions must be optimized to maximize therapeutic efficacy. These findings underscore the necessity of standardizing harvest and extraction protocols to ensure reproducibility and functional consistency of PDEVs for clinical applications. Overall, while classical isolation methods remain the foundation for PDEVs extraction, ongoing refinements and adaptations tailored to specific plant sources and research goals are critical to harness the full potential of plant-derived extracellular vesicles as drug delivery vehicles and therapeutic agents.32,51
PDEVs Biocompatibility and Immunogenicity
PDEVs have garnered significant attention as promising nanocarriers for targeted delivery of antitumor drugs, largely due to their intrinsic low immunogenicity and excellent biocompatibility. Unlike mammalian-derived extracellular vesicles (EVs), which often face challenges related to immune rejection and cytotoxicity, PDEVs are naturally endowed with properties that minimize immune system activation, making them suitable for long-term in vivo applications. Studies have demonstrated that PDEVs, isolated from various edible and medicinal plants, exhibit stable physicochemical characteristics and carry bioactive molecules such as proteins, lipids, microRNAs, and secondary metabolites that contribute to their therapeutic efficacy without eliciting adverse immune responses.22,52 The wide availability and cost-effectiveness of plant sources further enhance the appeal of PDEVs as safe drug delivery vehicles.
The low immunogenicity of PDEVs is attributed to their plant-origin lipid bilayer composition and the absence of mammalian antigenic proteins, which reduces recognition by the host immune system. The low immunogenicity of PDEVs is not simply characterized by an absence of immune response; rather, it is attributable to fundamental differences in the glycosylation patterns of their membrane proteins compared to those of mammals.53 Specifically, PDEVs lack terminal sialic acid residues and are enriched with high-mannose glycans as well as plant-specific xylose and fucoside structures.54 When these carbohydrate moieties are presented on intact nanovesicles, they are effectively recognized by mannose receptors on antigen-presenting cells and other C-type lectins, promoting endocytosis without activating downstream pro-inflammatory signaling pathways.53 This mechanism facilitates the natural targeting of intestinal immune cells upon oral administration of PDEVs, thereby inducing tolerogenic immune responses. This phenomenon contrasts with the traditionally held view that soluble plant glycoproteins primarily elicit sensitizing immune effects. For example, grapefruit-derived EVs loaded with therapeutic agents showed no hemolysis or organ toxicity in vascular calcification models, indicating their favorable safety profile.55 Similarly, Momordica charantia small extracellular vesicles demonstrated excellent biosafety upon intravenous administration in ischemic stroke models, with no significant systemic toxicity observed.56 These findings underscore the potential of PDEVs to circumvent immune clearance mechanisms that often limit the efficacy of synthetic or animal-derived nanocarriers. Moreover, PDEVs have been shown to modulate immune responses beneficially. Ginger-derived extracellular vesicles conjugated with folic acid selectively targeted inflammatory macrophages and promoted polarization towards reparative M2 phenotypes, thereby remodeling the immune microenvironment in rheumatoid arthritis without eliciting immunogenicity.57 This immunomodulatory capacity, combined with low immunogenicity, positions PDEVs as ideal candidates for chronic disease treatment requiring repeated or sustained administration. The biocompatibility of PDEVs extends to their ability to cross biological barriers efficiently, including the blood-brain barrier, facilitating drug delivery to otherwise inaccessible tissues.58 Their inherent stability in harsh physiological environments, such as the gastrointestinal tract, further supports their suitability for oral or mucosal administration routes, which are advantageous for patient compliance and reduced systemic side effects.59,60
Despite these promising attributes, challenges remain in standardizing isolation, characterization, and storage methods to ensure consistent PDEVs quality and functionality for clinical translation.30,61 Nonetheless, the accumulating preclinical evidence highlights that PDEVs possess a unique combination of low immunogenicity, high biocompatibility, and therapeutic versatility, making them highly suitable for long-term in vivo applications aimed at targeted antitumor drug delivery with minimized immune rejection risks.
Advances in the Application of PDEVs in Targeted Delivery of Antitumor Drugs
Drug Loading Strategies and Delivery Efficiency
PDEVs have emerged as promising natural nanocarriers for targeted drug delivery, owing to their intrinsic biocompatibility, stability, and ability to traverse biological barriers without eliciting significant immune responses or cytotoxicity.52,62 Efficient drug loading into PDEVs is critical to enhance therapeutic efficacy, improve drug stability, and ensure effective accumulation at tumor sites. Common drug loading strategies include co-incubation, chemical modification, electroporation, sonication, and extrusion, each with distinct advantages and limitations regarding loading efficiency, vesicle integrity, and scalability.63,64 Co-incubation, a simple and non-disruptive method, allows passive diffusion of hydrophobic drugs into the lipid bilayer of PDEVs, preserving vesicle structure and bioactivity.65 Chemical conjugation or surface modification can enhance targeting specificity and loading capacity, though these approaches require careful optimization to avoid compromising vesicle functionality.66 Advanced techniques such as light-triggered loading utilize photosensitizer-induced reactive oxygen species to transiently increase membrane permeability, thereby improving drug encapsulation without significant vesicle damage.67 Moreover, the physicochemical properties of both the drug and the PDEVs influence loading efficiency; for example, lipid solubility positively correlates with loading capacity, as demonstrated with ginger-derived EVs and gingerol compounds.68 The choice of plant source and isolation method also affects vesicle yield, composition, and drug-loading potential, with callus-derived vesicles showing variable loading efficiencies compared to apoplastic fluid-derived vesicles.69 Importantly, PDEVs can encapsulate both hydrophilic and hydrophobic agents, protecting them from degradation and enhancing bioavailability.70 For instance, curcumin-loaded tomato-derived nanovesicles exhibited improved anti-inflammatory activity compared to free curcumin, highlighting the functional synergy between the vesicle cargo and the plant-derived matrix.71 The oral administration route has been successfully explored using PDEVs, which can protect drugs from gastrointestinal degradation and facilitate targeted delivery to tumor sites.52 Collectively, these strategies underscore the versatility and efficacy of PDEVs as drug delivery vehicles, with ongoing research focused on optimizing loading techniques to maximize therapeutic payloads while maintaining vesicle stability and targeting capabilities.
A representative example illustrating the successful application of drug loading and targeted delivery via PDEVs involves extracellular vesicles derived from Isodon rubescens, a medicinal plant known for its bioactive compounds. In this case, vesicles were loaded with doxorubicin (DOX), a potent chemotherapeutic agent, through co-incubation or sonication methods to enhance drug encapsulation efficiency.66,69 These Isodon rubescens-derived vesicles facilitated oral administration of DOX, overcoming the common challenges of poor bioavailability and systemic toxicity associated with conventional chemotherapy. The loaded vesicles exhibited enhanced accumulation of DOX at tumor sites, as evidenced by in vivo biodistribution studies, leading to significant tumor growth inhibition with reduced off-target effects.52 Mechanistically, the vesicles’ natural lipid bilayer and surface proteins contributed to efficient cellular uptake and endosomal escape, ensuring effective intracellular drug release. Furthermore, functionalization of these vesicles with targeting ligands, such as antibodies against death receptor 5 (DR5), enabled selective delivery to senescent tumor cells, thereby overcoming therapy-induced senescence and suppressing the senescence-associated secretory phenotype that promotes tumor progression.66 This approach not only enhanced the cytotoxicity of DOX but also modulated the tumor immune microenvironment, demonstrating a multifaceted therapeutic benefit. The oral delivery of Isodon rubescens-derived vesicles loaded with DOX exemplifies the potential of PDEVs to serve as efficient, safe, and targeted drug delivery platforms, particularly for anticancer therapies. This strategy leverages the natural properties of plant vesicles to improve drug stability, bioavailability, and tumor specificity, offering a promising avenue for clinical translation in oncology.69,72
Mechanisms Targeting the TME
PDEVs have emerged as promising agents capable of modulating the TME through multiple mechanisms that collectively enhance antitumor efficacy. One critical aspect of PDEVs’ function is their ability to regulate the TME by inhibiting tumor immune escape, promoting tumor cell apoptosis, and improving drug efficacy. For example, Citrus limon-derived extracellular vesicles encapsulating temozolomide (TMZ) demonstrated enhanced drug stability and effective crossing of the blood-tumor barrier while preserving endothelial barrier integrity and reducing oxidative stress in glioblastoma models. Importantly, these vesicles suppressed vascular endothelial growth factor A (VEGF-A) secretion, thereby disrupting pro-tumorigenic angiogenic pathways within the TME.73 Similarly, Aloe vera-derived extracellular vesicle-like particles (EVLPs) inhibited pancreatic carcinoma progression by triggering mitochondrial reactive oxygen species (ROS) release and activating pyroptosis pathways, which led to increased tumor cell apoptosis and reduced tumor growth without systemic toxicity.74 These findings underscore the potential of PDEVs to directly induce tumor cell death and modulate the biochemical milieu of the TME to favor therapeutic outcomes.
Beyond direct tumor cell targeting, PDEVs also modulate the activity of tumor-associated immune cells, thereby enhancing antitumor immune responses. Curcuma-derived extracellular vesicles functionalized with antibodies targeting death receptor 5 (DR5) effectively eliminated senescent tumor cells and suppressed the senescence-associated secretory phenotype (SASP), which is known to create an immunosuppressive microenvironment. This approach not only promoted apoptosis of resistant tumor cells but also inhibited epithelial-mesenchymal transition and angiogenesis, resulting in a remodeled tumor immune microenvironment conducive to improved chemotherapy efficacy.66 Moreover, plant sap-derived extracellular vesicles from Dendropanax morbifera suppressed cancer-associated fibroblasts (CAFs), key mediators of tumor metastasis and TME remodeling, by altering gene expression related to growth factors and extracellular matrix components. This anti-CAF activity highlights how PDEVs can disrupt stromal support for tumors, thus attenuating metastatic potential.75
The immunomodulatory effects of PDEVs extend to the regulation of tumor-associated macrophages (TAMs), natural killer (NK) cells, dendritic cells, and T lymphocytes within the TME. Tumor-derived extracellular vesicles are known to facilitate immune evasion by transferring immunosuppressive molecules; however, PDEVs can counteract these effects by promoting immune cell activation and reprogramming the TME toward an antitumor phenotype. For instance, studies have shown that EVs can modulate the tumor immune microenvironment by altering the phenotypes and functions of various immune cells, thereby enhancing immune surveillance and cytotoxicity against tumor cells.76,77 The ability of PDEVs to deliver bioactive cargos such as microRNAs and proteins that regulate immune cell signaling pathways is a key mechanism by which they enhance antitumor immunity. For example, watermelon-derived EVs combined with dendrimer-bound miRNA146 mimic exhibited anti-angiogenic activity, which is critical for disrupting the immunosuppressive and nutrient-rich TME that supports tumor growth.28
Furthermore, PDEVs contribute to remodeling the extracellular matrix (ECM) and inhibiting tumor-promoting stromal cells, which indirectly supports immune-mediated tumor suppression. By altering the ECM composition and reducing the presence of CAFs, PDEVs can decrease physical and biochemical barriers to immune cell infiltration and drug delivery, thereby improving therapeutic efficacy.75,78 This multifaceted modulation of the TME by PDEVs not only impedes tumor progression but also sensitizes tumors to conventional therapies, as evidenced by meta-analyses showing that PDEVs combined with chemotherapeutic agents exhibit enhanced tumor volume reduction compared to free drugs alone.79 Moreover, through the genetic modification of plant organisms, the therapeutic capabilities of PDEVs can be expanded and tailored to specific needs. For instance, the human enzyme indoleamine 2,3-dioxygenase 1 (IDO1) has been successfully expressed in plant cells and targeted to MVBs, which serve as critical subcellular sites for PDEV biogenesis. This finding indicates that PDEVs can be engineered to encapsulate active IDO1 via this intracellular trafficking route.80 Upon oral administration, these vesicles are capable of locally producing kynurenine within the intestinal environment, thereby promoting the differentiation of regulatory T cells. This mechanism synergizes with the ECM remodeling properties inherent to natural PDEVs, facilitating the conversion of immunologically “cold” tumors into “hot” tumors while concurrently mitigating immune-related adverse effects.
In summary, PDEVs target the tumor microenvironment through a combination of direct tumor cell apoptosis induction, suppression of immunosuppressive phenotypes such as SASP, modulation of immune cell activity to enhance antitumor immunity, and remodeling of the stromal and extracellular matrix components (Figure 2). These mechanisms collectively inhibit tumor immune escape, promote tumor cell death, and improve the efficacy of anticancer drugs, positioning PDEVs as a promising biocompatible platform for targeted cancer therapy with potential for clinical translation.
 |
Figure 2 The mechanism of action of PDEVs in delivering anti-tumor drugs. PDEVs enhance anti-tumor immunity and reshape matrix and extracellular matrix components by directly inducing apoptosis of tumor cells and regulating the activity of immune cells. These mechanisms jointly inhibit tumor immune escape, promote tumor cell death, and enhance the efficacy of anti-cancer drugs. |
Combination Therapy and Multifunctional Carrier Design
Combination therapy involving PDEVs and conventional chemotherapeutic agents, nanomaterials, or fusion proteins represents a promising strategy to enhance therapeutic selectivity and anti-tumor efficacy. PDEVs serve as natural nanocarriers with inherent biocompatibility, low immunogenicity, and scalable production, making them ideal platforms for drug delivery. For instance, grapefruit-derived EVs have been demonstrated to effectively deliver chemotherapeutics such as paclitaxel, improving drug stability and bioavailability while reducing systemic toxicity in breast cancer models, including triple-negative breast cancer (TNBC).81 The integration of PDEVs with conventional chemotherapy agents can potentiate anti-tumor effects by facilitating targeted delivery and minimizing off-target damage. Furthermore, hybrid systems combining PDEVs with synthetic polymers or dendrimers have been developed to enhance cargo loading and delivery efficiency, as exemplified by watermelon-derived vesicles hybridized with polyamidoamine dendrimers loaded with miRNA mimics, which exhibited anti-angiogenic and anti-tumor activities.28 The design of multifunctional carriers also extends to the incorporation of stimuli-responsive elements, enabling controlled release and improved tumor targeting. For example, protein-based multifunctional delivery systems conjugated with targeting ligands such as cRGD peptides and membrane-translocating agents like protamine sulfate have demonstrated enhanced intracellular accumulation and inhibitory effects on tumor progression.82 Additionally, multifunctional nanoparticles co-loading chemotherapeutic drugs and photosensitizers have been engineered to combine chemotherapy with photothermal or sonodynamic therapy, achieving synergistic tumor suppression and reduced side effects.83,84 These multifunctional platforms often leverage natural biomaterials such as polysaccharides or marine-derived compounds to improve biocompatibility and therapeutic efficacy.85,86 Overall, the convergence of PDEVs with traditional chemotherapeutics, nanomaterials, and fusion proteins in multifunctional carrier systems holds great potential to overcome limitations of monotherapies, including multidrug resistance and systemic toxicity, by enhancing selective delivery, modulating tumor microenvironments, and enabling combination modalities.
In parallel, the integration of plant bioactive components such as bitter melon extract into these delivery platforms has been shown to augment cytotoxicity and modulate critical signaling pathways involved in tumorigenesis. Broccoli-derived extracellular vesicles enriched with selenium have been reported to carry miRNAs, such as miR167a, which induce apoptosis in pancreatic cancer cells by targeting IRS1 and interfering with the PI3K-AKT pathway.87 Similarly, plant-derived EVs from broccoli and other fruits transport functional miRNAs and phytochemicals that exert toxic effects on cancer cells by regulating gene expression and signaling cascades.88 Grapefruit-derived exosome-like nanovesicles containing antioxidant molecules like ascorbic acid have demonstrated selective antiproliferative effects on leukemic cells by increasing reactive oxygen species (ROS) levels, thereby enhancing the efficacy of existing therapies without harming normal cells.89 The combination of chemotherapeutic agents with plant-derived bioactive compounds encapsulated within PDEVs or multifunctional nanocarriers can synergistically enhance anti-tumor activity by simultaneously inducing cytotoxicity and modulating tumor-related signaling pathways. This dual approach not only improves therapeutic outcomes but also reduces adverse effects associated with high-dose chemotherapy. Moreover, the natural origin and multifunctionality of these carriers facilitate the remodeling of the tumor microenvironment and immune modulation, which are critical for sustained anti-cancer responses.79 Collectively, the strategic incorporation of plant bioactive components into PDEVs-based combination therapies and multifunctional carriers represents an innovative and effective avenue for advancing cancer treatment with improved selectivity, efficacy, and safety.
Therapeutic Application of PDEVs
EVs Derived From Momordica Charantia (Bitter Melon)
EVs derived from Momordica charantia (MCEVs) have emerged as potent bioactive nanovesicles exhibiting remarkable antitumor activities through multifaceted mechanisms. In vitro studies have demonstrated that MCEVs significantly inhibit cancer cell proliferation and induce apoptosis across various tumor cell lines, including cervical cancer cells such as HeLa and C33A. The half-maximal inhibitory concentrations (IC50) of MCEVs in these cells are approximately 59 µg/mL and 68 µg/mL, respectively, indicating substantial cytotoxicity. Mechanistically, MCEVs downregulate proliferative markers such as PCNA and Cyclin D1, thereby suppressing cell cycle progression and colony formation. They also inhibit cell migration by reducing epithelial-mesenchymal transition (EMT) markers including N-cadherin, vimentin, and matrix metalloproteinase 9, which are critical for tumor invasiveness and metastasis. Apoptosis induction by MCEVs is evidenced by increased apoptotic rates, elevated TUNEL-positive cells, a decreased Bcl-2/Bax ratio, and reduced phosphorylation of Akt, highlighting the involvement of the Bcl-2/Bax/p-Akt signaling pathway. Additionally, MCEVs promote ferroptosis, a regulated form of cell death characterized by lipid peroxidation, through upregulation of ACSL4 and transferrin receptor and downregulation of glutathione peroxidase 4 (GPX4), further contributing to antitumor efficacy. These findings underscore the ability of MCEVs to modulate multiple tumor-associated signaling cascades, including apoptosis, ferroptosis, and EMT pathways, thereby exerting comprehensive antitumor effects.4,90 Moreover, the antioxidant capacity of MCEVs, demonstrated by their ability to scavenge reactive oxygen species and protect cells from oxidative damage, may further contribute to their therapeutic potential by mitigating oxidative stress-related tumor progression.91 Collectively, these data establish MCEVs as promising natural nanotherapeutics capable of targeting diverse oncogenic pathways to inhibit tumor growth and progression.
The antitumor efficacy and immunomodulatory effects of Momordica charantia-derived extracellular vesicles have been substantiated in various animal models, highlighting their translational potential. In vivo studies using xenograft models of cervical cancer have demonstrated that intraperitoneal administration of MCEVs significantly suppresses tumor growth, as evidenced by reduced tumor volume and weight. Histological analyses reveal decreased expression of proliferation markers such as Ki-67 and EMT marker N-cadherin within tumor tissues, confirming the inhibitory effects on tumor cell proliferation and metastasis. Beyond direct tumor suppression, MCEVs exhibit notable immunomodulatory properties by remodeling the tumor microenvironment and enhancing host immune responses. These vesicles modulate oxidative stress and inflammatory pathways, contributing to an improved immune milieu that supports antitumor activity. Furthermore, MCEVs have been shown to synergize with conventional chemotherapeutic agents, such as 5-fluorouracil, by enhancing drug efficacy and overcoming chemoresistance mechanisms. For instance, bitter melon-derived EVs downregulate NLRP3 inflammasome activation, a pathway implicated in chemotherapy resistance, thereby sensitizing oral squamous cell carcinoma cells to 5-fluorouracil treatment and improving therapeutic outcomes. This synergy not only potentiates anticancer effects but also may reduce required chemotherapeutic dosages, potentially minimizing adverse effects. Additionally, MCEVs demonstrate protective effects against chemotherapy-induced toxicity, such as doxorubicin-induced cardiotoxicity, by stabilizing key proteins like p62 and activating antioxidant pathways (eg, Nrf2/HO-1), which further supports their role as adjunctive agents in cancer therapy. The safety profile of MCEVs is favorable, with intravenous administration showing excellent biocompatibility in animal models. Collectively, these preclinical findings validate the dual antitumor and immunoregulatory functions of Momordica charantia-derived EVs and underscore their promise as natural nanocarriers to enhance chemotherapy efficacy and reduce treatment-associated toxicity in cancer management.4,90,92,93
EVs Derived From Isodon Rubescens
EVs derived from Isodon rubescens, a medicinal plant known for its potent bioactive compounds, have garnered significant attention due to their intrinsic anti-inflammatory and antitumor activities. These vesicles naturally encapsulate a variety of phytochemicals that modulate cellular signaling pathways involved in tumor progression and inflammation, rendering them promising candidates for cancer therapy. The inherent bioactivity of Isodon rubescens-derived EVs provides a dual advantage: they not only exert direct cytotoxic effects on malignant cells but also modulate the TME to suppress cancer-promoting inflammation. This unique property makes them particularly suitable as carriers for chemotherapeutic agents, enabling targeted delivery that enhances drug efficacy while minimizing systemic toxicity. By leveraging the natural affinity of these vesicles for tumor cells and their ability to penetrate biological barriers, chemotherapeutic drugs loaded into Isodon rubescens EVs can achieve higher local concentrations within tumors, thereby improving therapeutic outcomes. Moreover, the biocompatibility and low immunogenicity of plant-derived EVs reduce the risk of adverse immune reactions, which is a critical consideration in cancer treatment. The integration of Isodon rubescens EVs as delivery vehicles thus represents a promising strategy to harness both the plant’s natural pharmacological effects and the precision of nanomedicine for targeted anticancer therapy.94
To overcome the challenges associated with oral administration of anticancer agents, such as poor bioavailability and degradation in the gastrointestinal tract, chemical modification of Isodon rubescens-derived EVs has been employed to enhance their stability and targeting capabilities. These modifications may include surface functionalization with ligands that recognize receptors overexpressed in colon cancer cells or the incorporation of protective coatings that shield the vesicles from enzymatic degradation. Such engineered EVs demonstrate improved oral delivery efficiency, facilitating the transport of chemotherapeutic drugs across the intestinal mucosa and ensuring their accumulation at tumor sites within the colon. This targeted delivery approach not only maximizes the therapeutic index of anticancer drugs but also reduces off-target effects and systemic toxicity. Preclinical studies have shown that chemically modified Isodon rubescens EVs loaded with chemotherapeutic agents significantly inhibit tumor growth in colon cancer models, highlighting their potential to improve clinical outcomes in colon-related malignancies. Additionally, these vesicles can modulate the tumor microenvironment by delivering bioactive molecules that suppress inflammation and angiogenesis, further contributing to their antitumor efficacy. The ability to administer these EVs orally also enhances patient compliance and broadens their clinical applicability. Collectively, the chemical modification of Isodon rubescens-derived EVs represents a critical advancement in the development of plant-based nanocarriers for effective, targeted, and patient-friendly cancer therapies.94
Exploration of the Potential of Other PDEVs
PDEVs exhibit remarkable heterogeneity in their compositional and functional profiles depending on their botanical origin, which underpins their potential for developing diversified and tailored antitumor therapeutic strategies. This diversity arises from the intrinsic biochemical makeup of the source plants, which influences the vesicles’ cargo of lipids, proteins, nucleic acids, and bioactive phytochemicals, thereby modulating their biological activities and therapeutic efficacies. For instance, ginger-derived EVs (GDEVs) have been shown to contain cytotoxic compounds such as gingerols and shogaols, which contribute to their potent tumor cell inhibitory effects by inducing apoptosis through regulation of cell cycle and p53 signaling pathways.95 These GDEVs also demonstrate superior antioxidant and anti-inflammatory properties, making them effective in mitigating oxidative stress-related pathologies, including cancer.68,96 Similarly, extracellular vesicles isolated from Aloe vera (AV-EVLPs) display significant antitumor activities against pancreatic carcinoma by triggering pyroptosis via mitochondrial reactive oxygen species (ROS)-mediated caspase-GSDMD/E signaling pathways, highlighting their unique mechanistic role in tumor suppression.74
Furthermore, PDEVs derived from diverse fruits such as oranges, apples, and sea buckthorn reveal distinct physicochemical properties and bioactive profiles that influence their drug delivery capabilities and therapeutic outcomes. Orange-derived EVs differ in vesicle concentration, lipid composition, and protein content across varieties and maturity stages, which affects their capacity to encapsulate and stabilize polyphenols like curcumin, thereby enhancing bioavailability and antioxidant activity.50,70 Apple-derived vesicles have demonstrated safety and efficacy in dermatological applications, providing anti-aging and soothing benefits through their bioactive cargo.97 Sea buckthorn EVs promote bone regeneration via miRNA-mediated pathways, underscoring the potential of PDEVs in regenerative oncology and tissue repair.98
Proteomic and metabolomic analyses further accentuate the compositional distinctions among PDEVs from different plant tissues and species. For example, potato root and peel-derived EVs show significant differences in size and protein enrichment, with unique proteins implicated in biotic and abiotic stress responses, suggesting differential roles in plant defense and potential therapeutic applications.99 Similarly, EVs from medicinal plants such as Drynariae Rhizoma roots are enriched with enzymes involved in oxidative phosphorylation pathways linked to neurodegenerative diseases, indicating their multifaceted therapeutic potential beyond oncology.21 Cannabis strain-derived PDEVs exhibit variability in protein and RNA content dependent on cultivar and age, which may influence their biomarker and therapeutic utility.100
The functional versatility of PDEVs is also evident in their capacity to modulate immune responses, inflammation, and the tumor microenvironment. Studies have demonstrated that PDEVs can regulate immune cell behavior, promote anti-inflammatory cytokine production, and restore tissue homeostasis, which are critical factors in cancer progression and therapy resistance.101,102 Moreover, the ability of PDEVs to cross biological barriers and their inherent low immunogenicity make them attractive natural nanocarriers for targeted drug delivery, enhancing the precision and efficacy of antitumor agents while minimizing off-target effects.1,33 The engineering of PDEVs, including surface modifications and drug loading techniques, further expands their applicability in personalized cancer therapy.72,103
In conclusion, the compositional and functional heterogeneity of PDEVs derived from different plant species and tissues offers a rich resource for the development of multifaceted and customizable tumor treatment strategies. Harnessing this diversity through systematic characterization and optimization of isolation methods will enable the design of PDEVs-based therapeutics that exploit specific bioactive cargos and targeting capabilities. This approach holds promise for advancing precision oncology by integrating natural nanocarriers with intrinsic therapeutic properties, thereby enhancing treatment efficacy and safety profiles in preclinical and clinical settings.32,104
Applications and Challenges of PDEVs in Preclinical Studies
Preclinical Animal Model Research Progress
Preclinical animal studies have provided compelling evidence that PDEVs loaded with anticancer drugs can effectively deliver therapeutic agents in vivo, resulting in significant tumor growth inhibition while concurrently reducing systemic toxicity. These in vivo experiments highlight the unique advantages of PDEVs as delivery vehicles, including their biocompatibility, ability to cross biological barriers, and inherent targeting capabilities. Engineered EVs, including those derived from plants, have been demonstrated to mediate precise delivery of chemotherapeutic compounds, antibodies, and nucleic acid-based therapeutics such as siRNA and CRISPR/Cas9 systems directly to tumor sites, thereby enhancing antitumor efficacy and limiting off-target effects.94 The multifunctional nature of these vesicles allows them not only to suppress malignant phenotypes of cancer cells but also to modulate the TME, eliciting antitumor immune responses and inhibiting tumor angiogenesis, which are critical factors in tumor progression and metastasis. Various animal models, including murine xenograft and orthotopic tumor models, have validated the safety profile of PDEVs-based drug delivery systems, demonstrating minimal immunogenicity and systemic toxicity compared to conventional chemotherapy. These studies collectively underscore the therapeutic potential of PDEVs in cancer treatment by confirming their ability to achieve targeted drug delivery, potentiate antitumor activity, and reduce adverse side effects in vivo. The translational relevance of these findings suggests that PDEVs could serve as a versatile platform for the development of next-generation anticancer therapies with improved efficacy and safety profiles, paving the way for future clinical applications.94
Pharmacokinetics and Biodistribution Characteristics
PDEVs have emerged as promising nanocarriers that significantly enhance the pharmacokinetic profiles and biodistribution of antitumor drugs, addressing critical limitations such as poor stability, rapid clearance, and nonspecific tissue distribution. One key advantage of PDEVs is their ability to improve drug stability in the systemic circulation. For instance, encapsulation of bioactive compounds like the plant saponin Uttroside B (Utt-B) within extracellular vesicles derived from Solanum nigrum Linn leaves demonstrated enhanced cellular uptake and sustained release kinetics, as confirmed by physicochemical characterization and pharmacokinetic simulations using physiologically based pharmacokinetic (PBPK) modeling. This encapsulation not only protected Utt-B from premature degradation but also improved its bioavailability upon administration, indicating a prolonged half-life compared to free drug formulations.105 Similarly, PDEVs derived from grapefruit juice have been shown to efficiently deliver exogenous proteins such as bovine serum albumin and heat shock protein 70 into human cells, with in vivo biodistribution studies revealing widespread organ uptake and suggesting effective systemic delivery with minimal toxicity.106
Moreover, PDEVs exhibit inherent biocompatibility and low immunogenicity, which contribute to their favorable pharmacokinetics by reducing rapid clearance by the mononuclear phagocyte system (MPS). Positron emission tomography (PET) imaging studies of extracellular vesicles from various human cell sources have highlighted that EVs generally display extended circulation times and relatively low accumulation in clearance organs such as liver and spleen, although biodistribution can vary depending on the cell source and labeling method. This suggests that PDEVs, with their distinct lipid and protein compositions, can be engineered or selected to optimize circulation time and tissue targeting.107 The selective targeting capability of PDEVs is further supported by their ability to remodel the TME and enhance antitumor immunity, which is crucial for precision oncology. For example, plant-derived EVs have been implicated in modulating immune responses and overcoming tumor immune evasion, thereby improving the localization and retention of therapeutic agents at tumor sites.1
The pharmacokinetic improvements afforded by PDEVs also translate into enhanced therapeutic efficacy with reduced off-target effects. In glioblastoma models, extracellular vesicles derived from bone marrow mesenchymal stem cells (BMSC-EVs) loaded with the anti-cancer drug celastrol exhibited superior brain tumor targeting, facilitated blood-brain barrier penetration, and increased tumor apoptosis compared to free drug administration, while simultaneously decreasing systemic toxicity. This demonstrates the ability of EVs to prolong drug half-life and concentrate therapeutic payloads within tumors, minimizing damage to healthy tissues.108 Additionally, plant-based exosome-like vesicles have shown promise in breast cancer therapy by improving drug bioavailability and stability, which reduces systemic adverse effects and enhances drug accumulation at tumor sites.109
Furthermore, the intrinsic properties of PDEVs, such as their lipid bilayer composition and surface molecules, contribute to their efficient cellular uptake and targeted delivery. Studies have demonstrated that PDEVs can enter cells via mechanisms like macropinocytosis, facilitating intracellular delivery of cargo molecules and enabling sustained pharmacological action.98 This targeted delivery is critical for prolonging the half-life of drugs within the tumor microenvironment, thereby enhancing antitumor efficacy while sparing non-target tissues.
In summary, PDEVs improve the pharmacokinetic and biodistribution profiles of antitumor drugs by enhancing drug stability in circulation, prolonging half-life, and enabling targeted delivery to tumor tissues. These properties collectively reduce off-target toxicity and improve therapeutic outcomes. The biocompatibility, natural origin, and modifiable surface characteristics of PDEVs position them as a highly promising platform for the development of next-generation targeted anticancer drug delivery systems with superior pharmacokinetic behavior and clinical potential. Continued research into optimizing PDEVs isolation, drug loading, and targeting strategies will further advance their application in precision oncology.
Technical and Translational Challenges
The clinical translation of PDEVs as targeted delivery systems for antitumor drugs faces significant technical and translational hurdles, notably in standardization, quality control, drug loading efficiency, release mechanisms, in vivo metabolism, and immunogenicity. Herein, we will discuss the development and critical issues associated with the scale-up production and clinical research of PDEVs (Figure 3). Standardized extraction and purification protocols remain elusive due to the inherent complexity and variability of plant sources and the diverse physicochemical properties of PDEVs. Various isolation techniques such as ultracentrifugation, size-exclusion chromatography, and precipitation methods have been employed, but lack of consensus on optimal protocols leads to batch-to-batch variability and challenges in reproducibility.61,110 This variability complicates the establishment of quality control criteria essential for clinical-grade PDEVs, including particle size distribution, surface charge, molecular cargo characterization, and functional potency. Moreover, the drug loading efficiency into PDEVs and the control over drug release kinetics require further optimization. Current studies show that PDEVs can encapsulate both hydrophilic and hydrophobic drugs effectively, as demonstrated with compounds like curcumin and doxorubicin, yet the mechanisms governing drug encapsulation, retention, and controlled release remain incompletely understood.111,112 Enhancing loading strategies and engineering surface modifications are active research areas aimed at improving targeting specificity and therapeutic efficacy.62
|
Figure 3 The development and critical issues of PDEVs-based nano-drug delivery can be subdivided into upstream and downstream processing and preclinical study. |
In vivo metabolic pathways of PDEVs also present a critical knowledge gap. While PDEVs have demonstrated the ability to cross biological barriers such as the gastrointestinal tract and blood-brain barrier, their biodistribution, clearance mechanisms, and potential accumulation in off-target tissues require comprehensive investigation to ensure safety and efficacy.5,113 The immunogenic profile of PDEVs is generally favorable, exhibiting low immunogenicity compared to mammalian-derived vesicles, which is advantageous for repeated administration. Nonetheless, potential immune responses, especially in the context of repeated dosing or in patients with altered immune status, must be rigorously evaluated in preclinical and clinical studies.103,114 Furthermore, the influence of plant source variability, including differences in vesicle composition and bioactive cargo, on immunogenicity and therapeutic outcomes is not fully elucidated.70
Translational challenges also encompass scalability and manufacturing under good manufacturing practice (GMP) conditions. The scalability of PDEVs production is promising due to the abundance and renewability of plant materials; however, establishing robust, reproducible, and cost-effective large-scale production processes remains a bottleneck.115,116 Storage stability is another critical factor; PDEVs must retain their structural integrity and bioactivity during storage and transport, which is complicated by their lipid bilayer composition susceptible to degradation.117 Strategies such as lyophilization and incorporation into hydrogels have been explored to enhance stability but require further validation.111
In summary, overcoming the technical challenges of standardized extraction, quality control, optimized drug loading and release, and understanding in vivo metabolism and immunogenicity are essential steps toward clinical translation of PDEVs as antitumor drug delivery vehicles. Addressing these issues through interdisciplinary research integrating plant biology, nanotechnology, pharmacology, and clinical sciences will facilitate the development of safe, effective, and scalable PDEVs-based therapeutics with significant potential to improve cancer treatment outcomes.1,118
Future Prospects: Application Prospects of PDEVs in Precision Tumor Therapy
PDEVs are emerging as versatile platforms for antitumor therapy, uniquely combining intrinsic biocompatibility, low immunogenicity, and the ability to traverse biological barriers with extensive engineering potential.1 Multifunctional intelligent carriers based on PDEVs integrate targeting ligands (antibodies, peptides, aptamers) and stimuli-responsive elements (pH, redox, enzymes, light) to achieve spatiotemporal control of drug release within the tumor microenvironment (TME). Chemical functionalization, such as deoxycholic acid modification of Isodon rubescens-derived vesicles, enables targeted oral delivery of doxorubicin to colon tumors with minimal systemic toxicity.5 Loading recombinant HSP70 into grapefruit-derived vesicles enhances antitumor immunity at significantly lower doses than free protein, highlighting the advantage of encapsulation. Harnessing endogenous TME stimuli, Aloe vera-derived vesicles induce pyroptosis via ROS-mediated pathways, while the natural cargo of PDEVs (lipids, proteins, nucleic acids) synergizes with chemotherapeutics, as demonstrated by bitter melon-derived vesicles enhancing 5-fluorouracil efficacy and reducing drug resistance in oral squamous cell carcinoma.92
In combination immunotherapy, PDEVs reprogram the TME by promoting M1-like macrophage polarization and dendritic cell maturation, thereby stimulating cytotoxic T lymphocyte responses and sensitizing tumors to immune checkpoint inhibitors (ICIs).119,120 PDEVs act as adjuvants or delivery vehicles for immunostimulatory molecules, siRNAs targeting immunosuppressive pathways, or tumor-associated antigens, synergizing with ICIs to alleviate immune suppression and reinvigorate exhausted T cells. Their ability to modulate gut microbiota offers an additional indirect mechanism to improve immunotherapy outcomes.
Clinical translation and industrialization require standardized, GMP-compliant production workflows, including robust isolation methods (ultracentrifugation, precipitation) and comprehensive characterization (size, surface markers, cargo, functional assays) to ensure batch-to-batch consistency.61 Safety evaluation must address immunogenicity, toxicity, biodistribution, and pharmacokinetics.73 Interdisciplinary collaboration-integrating plant biology, biotechnology, nanomedicine, and clinical research-is essential to optimize plant sources, engineering strategies, and translational study design.
A frontier direction involves genetically engineering plants as bioreactors to produce therapeutic vesicles. Expression of human indoleamine 2,3-dioxygenase 1 (IDO1) in tobacco cells yields active enzyme that localizes to multivesicular bodies, the key subcellular compartment for PDEV biogenesis. Such engineered vesicles could deliver IDO1 orally to generate kynurenine locally, inducing regulatory T cells and synergizing with the inherent antitumor activities of PDEVs. This approach expands the therapeutic repertoire to include immunomodulatory enzymes, cytokines, tumor antigens, and RNA drugs, leveraging endogenous plant sorting pathways for efficient loading. Future efforts must address heterologous protein stability, the immunomodulatory impact of plant-specific glycosylation, and the development of tailored regulatory frameworks to realize the full clinical potential of engineered PDEVs.
Conclusion
In conclusion, PDEVs have emerged as a promising class of natural nanocarriers for targeted antitumor drug delivery, offering distinct advantages that address several limitations of conventional delivery systems. Despite encouraging advances in preclinical studies demonstrating the feasibility and therapeutic benefits of PDEVs, several critical challenges remain before their widespread clinical application can be realized. Standardizing extraction and purification protocols is essential to ensure reproducibility, scalability, and regulatory compliance. Additionally, comprehensive pharmacokinetic profiling and rigorous safety assessments are imperative to elucidate their biodistribution, metabolism, and potential off-target effects. Through continuous innovation, rigorous evaluation, and strategic collaboration, these natural nanocarriers may potentially become safe, effective and multi-functional anti-cancer tools in the near future.
Author Contributions
Defa Huang and Jie Chen contributed equally to this work and share first authorship. All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the Ganzhou Science and Technology Plan Project (2023LNS37081 and GZ2024YLJ137), Jiangxi Provincial Health Commission Technology Plan Project (202510463), Jiangxi Province Administration of Traditional Chinese Medicine Scienceand Technology Plan Project (2024B0303) and Science and Technology Research Project of Jiangxi Provincial Department of Education (GJJ2501317).
Disclosure
The authors declare no conflict of interest. All authors agreed and reviewed the manuscript.
References
1. Wu H, Shi M, Meng L, et al. Plant-derived extracellular vesicles as a novel tumor-targeting delivery system for cancer treatment. Front Cell Dev Biol. 2025;13:1589550. doi:10.3389/fcell.2025.1589550
2. Gao J, Wu Y, Zhao Y, Lu Y, Cai X. Application of heterologous extracellular vesicles and their analogs in anti-tumor activity research. Colloids Surf B. 2025;254:114850. doi:10.1016/j.colsurfb.2025.114850
3. Maricchiolo E, Creanza P, Osnato M, et al. Plant vs mammal extracellular vesicles: new tools in therapeutic drug delivery. Current Res Biotechnol. 2026;11:100352. doi:10.1016/j.crbiot.2025.100352
4. Deligiannidou GE, Pritsa A, Nikolaou A, et al. Nutraceutical potential of bitter melon (Momordica charantia) on cancer treatment: an overview of in vitro and animal studies. Current Issues Mol Biol. 2025;47:425. doi:10.3390/cimb47060425
5. Zhang J, Zhang Y, Ma X, Wang H, Zhang C, Pan C. Screening Isodon rubescens-derived vesicles harvested at different periods as carriers for the treatment of colitis-associated cancer. ACS Biomater Sci Eng. 2025;11:2653–19. doi:10.1021/acsbiomaterials.5c00160
6. Hussain Z, Nigri J, Tomasini R. The cellular and biological impact of extracellular vesicles in pancreatic cancer. Cancers. 2021;13(12):3040. doi:10.3390/cancers13123040
7. Zhou Y, Liu M, Lu N, et al. Advances in the design and application of glycolysis-related nanodelivery systems for anti-tumor. Colloids Surf B. 2025;254:114844. doi:10.1016/j.colsurfb.2025.114844
8. Ma W, Wang X, Zhang D, Mu X. Research progress of disulfide bond based tumor microenvironment targeted drug delivery system. Int J Nanomed. 2024;19:7547–7566. doi:10.2147/IJN.S471734
9. Liu Y, Zhang Z, Du H, et al. An enzyme-responsive hydrogel functionalized with mesoporous silica nanoparticles for co-delivery of cisplatin and shRNA to overcome chemotherapy resistance in non-small cell lung cancer. RSC Adv. 2025;15:23966–23977. doi:10.1039/D5RA03250D
10. Guo R, Wang P. Tumor-derived extracellular vesicles: Hijacking T cell function through exhaustion. Pathol Res Pract. 2025;269:155948. doi:10.1016/j.prp.2025.155948
11. Wang S, Sun J, Dastgheyb RM, Li Z. Tumor-derived extracellular vesicles modulate innate immune responses to affect tumor progression. Front Immunol. 2022;13:1045624. doi:10.3389/fimmu.2022.1045624
12. Huang H, Wang X, Gao Z, et al. A platelet-powered drug delivery system for enhancing chemotherapy efficacy for liver cancer using the trojan horse strategy. Pharmaceutics. 2024;16(7):905. doi:10.3390/pharmaceutics16070905
13. Han R, Liu Q, Lu Y, et al. Tumor microenvironment-responsive Ag(2)S-PAsp(DOX)-cRGD nanoparticles-mediated photochemotherapy enhances the immune response to tumor therapy. Biomaterials. 2022;281:121328. doi:10.1016/j.biomaterials.2021.121328
14. An Q, van Bel AJ, Huckelhoven R. Do plant cells secrete exosomes derived from multivesicular bodies? Plant Signal Behav. 2007;2:4–7. doi:10.4161/psb.2.1.3596
15. Cai Q, He B, Jin H. A safe ride in extracellular vesicles – small RNA trafficking between plant hosts and pathogens. Curr Opin Plant Biol. 2019;52:140–148. doi:10.1016/j.pbi.2019.09.001
16. Nemati M, Singh B, Mir RA, et al. Plant-derived extracellular vesicles: a novel nanomedicine approach with advantages and challenges. Cell Commun Signal. 2022;20:69. doi:10.1186/s12964-022-00889-1
17. Movahed N, Cabanillas DG, Wan J, Vali H, Laliberté JF, Zheng H. Turnip mosaic virus components are released into the extracellular space by vesicles in infected leaves. Plant Physiol. 2019;180:1375–1388. doi:10.1104/pp.19.00381
18. Jin Z, Na J, Lin X, Jiao R, Liu X, Huang Y. Plant-derived exosome-like nanovesicles: a novel nanotool for disease therapy. Heliyon. 2024;10:e30630. doi:10.1016/j.heliyon.2024.e30630
19. An Q, Ehlers K, Kogel KH, van Bel AJ, Hückelhoven R. Multivesicular compartments proliferate in susceptible and resistant MLA12-barley leaves in response to infection by the biotrophic powdery mildew fungus. New Phytol. 2006;172:563–576. doi:10.1111/j.1469-8137.2006.01844.x
20. Li B, Jin Y, Xu Y, Wu Y, Xu J, Tu Y. Safety evaluation of tea (Camellia sinensis (L.) O. Kuntze) flower extract: assessment of mutagenicity, and acute and subchronic toxicity in rats. J Ethnopharmacol. 2011;133:583–590. doi:10.1016/j.jep.2010.02.030
21. Cao Y, Zhao Q, Liu F, et al. Drug value of Drynariae Rhizoma root-derived extracellular vesicles for neurodegenerative diseases based on proteomics and bioinformatics. Plant Signal Behav. 2022;17:2129290. doi:10.1080/15592324.2022.2129290
22. Feng J, Xiu Q, Huang Y, Troyer Z, Li B, Zheng L. Plant-derived vesicle-like nanoparticles as promising biotherapeutic tools: present and future. Adv Mater. 2023;35:e2207826. doi:10.1002/adma.202207826
23. Isaac R, Reis FCG, Ying W, Olefsky JM. Exosomes as mediators of intercellular crosstalk in metabolism. Cell Metab. 2021;33:1744–1762. doi:10.1016/j.cmet.2021.08.006
24. Hatsugai N, Iwasaki S, Tamura K, et al. A novel membrane fusion-mediated plant immunity against bacterial pathogens. Genes Dev. 2009;23:2496–2506. doi:10.1101/gad.1825209
25. Dad HA, Gu TW, Zhu AQ, Huang LQ, Peng LH. Plant exosome-like nanovesicles: emerging therapeutics and drug delivery nanoplatforms. Mol Ther. 2021;29:13–31. doi:10.1016/j.ymthe.2020.11.030
26. Cong M, Tan S, Li S, et al. Technology insight: Plant-derived vesicles-How far from the clinical biotherapeutics and therapeutic drug carriers? Adv Drug Deliv Rev. 2022;182:114108. doi:10.1016/j.addr.2021.114108
27. Kırbaş OK, Sağraç D, Çiftçi Ö C, et al. Unveiling the potential: extracellular vesicles from plant cell suspension cultures as a promising source. BioFactors. 2025;51:e2090. doi:10.1002/biof.2090
28. Corvigno S, Liu Y, Bayraktar E, et al. Enhanced plant-derived vesicles for nucleotide delivery for cancer therapy. NPJ Precis Oncol. 2024;8:86. doi:10.1038/s41698-024-00556-3
29. Tiwari A, Soni N, Dongre S, Chaudhary M, Bissa B. The role of plant-derived extracellular vesicles in ameliorating chronic diseases. Mol Biol Rep. 2025;52:360. doi:10.1007/s11033-025-10466-7
30. Lv L, Li Z, Liu X, et al. Revolutionizing medicine: harnessing plant-derived vesicles for therapy and drug transport. Heliyon. 2024;10:e40127. doi:10.1016/j.heliyon.2024.e40127
31. Kathait P, Patel PK, Sahu AN. Harnessing exosomes and plant-derived exosomes as nanocarriers for the efficient delivery of plant bioactives. Nanomedicine. 2024;19:2679–2697. doi:10.1080/17435889.2024.2354159
32. Huang D, Chen J, Zhao M, et al. Plant-derived extracellular vesicles: composition, function and clinical potential. J Transl Med. 2025;23:1065. doi:10.1186/s12967-025-07101-1
33. Yang J, Ai X, Zhang C, Guo T, Feng N. Application of plant-derived extracellular vesicles as novel carriers in drug delivery systems: a review. Expert Opin Drug Delivery. 2025;22:787–803. doi:10.1080/17425247.2025.2487589
34. Hwang D, Vinod N, Skoczen SL, et al. Bioequivalence assessment of high-capacity polymeric micelle nanoformulation of paclitaxel and Abraxane® in rodent and non-human primate models using a stable isotope tracer assay. Biomaterials. 2021;278:121140. doi:10.1016/j.biomaterials.2021.121140
35. Fujita D, Arai T, Komori H, et al. Apple-derived nanoparticles modulate expression of organic-anion-transporting Polypeptide (OATP) 2B1 in Caco-2 cells. Mol Pharm. 2018;15:5772–5780. doi:10.1021/acs.molpharmaceut.8b00921
36. Wang Y, Wang J, Ma J, Zhou Y, Lu R. Focusing on future applications and current challenges of plant derived extracellular vesicles. Pharmaceuticals. 2022;15. doi:10.3390/ph15060708
37. Cao M, Yan H, Han X, et al. Ginseng-derived nanoparticles alter macrophage polarization to inhibit melanoma growth. J Immunother Cancer. 2019;7:326. doi:10.1186/s40425-019-0817-4
38. Lee R, Ko HJ, Kim K, et al. Anti-melanogenic effects of extracellular vesicles derived from plant leaves and stems in mouse melanoma cells and human healthy skin. J Extracell Vesicles. 2020;9:1703480. doi:10.1080/20013078.2019.1703480
39. Chen A, He B, Jin H. Isolation of extracellular vesicles from Arabidopsis. Curr Protoc. 2022;2:e352. doi:10.1002/cpz1.352
40. Rutter BD, Innes RW. Extracellular vesicles isolated from the leaf apoplast carry stress-response proteins. Plant Physiol. 2017;173:728–741. doi:10.1104/pp.16.01253
41. De Palma M, Ambrosone A, Leone A, et al. Plant roots release small extracellular vesicles with antifungal activity. Plants. 2020;9:1777.
42. Kim T, Hong JW, Lee LP. Efficient methods of isolation and purification of extracellular vesicles. Nano Convergence. 2025;12:45. doi:10.1186/s40580-025-00509-x
43. Li B, Wu W, Xu W, Qian H, Ji C. Advances of extracellular vesicles isolation and detection frontier technology: from heterogeneity analysis to clinical application. J Nanobiotechnol. 2025;23:678. doi:10.1186/s12951-025-03768-2
44. Huang Y, Wang S, Cai Q, Jin H. Effective methods for isolation and purification of extracellular vesicles from plants. J Integr Plant Biol. 2021;63:2020–2030. doi:10.1111/jipb.13181
45. Wang Q, Jing R, Cao N, et al. Generalizability of enzyme-based isolation approach for extracellular vesicles from traditional medicinal plants. J Extracell Biol. 2025;4:e70090. doi:10.1002/jex2.70090
46. Vestuto V, Conte M, Vietri M, et al. Multiomic profiling and neuroprotective bioactivity of salvia hairy root-derived extracellular vesicles in a cellular model of parkinson’s disease. Int J Nanomed. 2024;19:9373–9393. doi:10.2147/IJN.S479959
47. Conte M, Cappetta E, Alfieri M, et al. Isolation and biophysical characterization of extracellular vesicles from hairy root cultures. Biol Protoc. 2025;15:e5225. doi:10.21769/BioProtoc.5225
48. Kocholata M, Prusova M, Malinska HA, Maly J, Janouskova O. Comparison of two isolation methods of tobacco-derived extracellular vesicles, their characterization and uptake by plant and rat cells. Sci Rep. 2022;12:19896. doi:10.1038/s41598-022-23961-9
49. Wen Z, Yu J, Jeong H, et al. An all-in-one platform to deplete pathogenic bacteria for rapid and safe enrichment of plant-derived extracellular vesicles. Lab Chip. 2023;23:4483–4492. doi:10.1039/D3LC00585B
50. Liu H, Yang C, Wu X, et al. Influence of the maturity on the characteristics of Orange-derived extracellular vesicles and their delivery performance for curcumin. Food Chem. 2025;485:144518. doi:10.1016/j.foodchem.2025.144518
51. Zeng YB, Deng X, Shen LS, et al. Advances in plant-derived extracellular vesicles: isolation, composition, and biological functions. Food Funct. 2024;15:11319–11341. doi:10.1039/D4FO04321A
52. Langellotto MD, Rassu G, Serri C, Demartis S, Giunchedi P, Gavini E. Plant-derived extracellular vesicles: a synergetic combination of a drug delivery system and a source of natural bioactive compounds. Drug Delivery Transl Res. 2025;15:831–845. doi:10.1007/s13346-024-01698-4
53. Song Y, Feng N, Yu Q, et al. Exosomes in disease therapy: plant-derived exosome-like nanoparticles current status, challenges, and future prospects. Int J Nanomed. 2025;20:10613–10644. doi:10.2147/IJN.S540094
54. Wang Q, Du W, Wang H, et al. Nicotine’s effect on cognition, a friend or foe? Prog Neuro-Psychopharmacol Biol Psych. 2023;124:110723. doi:10.1016/j.pnpbp.2023.110723
55. Feng W, Teng Y, Zhong Q, et al. Biomimetic grapefruit-derived extracellular vesicles for safe and targeted delivery of sodium thiosulfate against vascular calcification. ACS Nano. 2023;17:24773–24789. doi:10.1021/acsnano.3c05261
56. Huang LY, Liu YN, Li LL, et al. Momordica charantia small extracellular vesicles mitigate neuronal ferroptosis by inhibition of GPX4 ubiquitination in ischemic stroke. Phytomedicine. 2025;148:157298. doi:10.1016/j.phymed.2025.157298
57. Han R, Zhou D, Ji N, et al. Folic acid-modified ginger-derived extracellular vesicles for targeted treatment of rheumatoid arthritis by remodeling immune microenvironment via the PI3K-AKT pathway. J Nanobiotechnol. 2025;23:41. doi:10.1186/s12951-025-03096-5
58. Yu Y, Xu Z, Xu L, Lu D, Tang Y, Mai H. Plant extracellular vesicles as emerging neuroprotective agents for central nervous system disorders. J Adv Res. 2025;2025:1.
59. Pomatto MAC, Gai C, Negro F, et al. Plant-derived extracellular vesicles as a delivery platform for RNA-based vaccine: feasibility study of an oral and intranasal SARS-CoV-2 vaccine. Pharmaceutics. 2023;15. doi:10.3390/pharmaceutics15030974
60. Pomatto MAC, Gai C, Negro F, et al. Oral delivery of mRNA vaccine by plant-derived extracellular vesicle carriers. Cells. 2023;12:1826. doi:10.3390/cells12141826
61. Kürtösi B, Kazsoki A, Zelkó R. A systematic review on plant-derived extracellular vesicles as drug delivery systems. Int J Mol Sci. 2024;25:7559. doi:10.3390/ijms25147559
62. Rajasekaran B, Lo KJ, Pan MH. Recent advances in PDEVs as nanocarriers for drug delivery: loading techniques, engineering strategies and future directions. Expert Opin Drug Delivery. 2025;23(1):145–168.
63. Man F, Wang J, Lu R. Techniques and applications of animal- and plant-derived exosome-based drug delivery system. J Biomed Nanotechnol. 2020;16:1543–1569. doi:10.1166/jbn.2020.2993
64. Zeng H, Guo S, Ren X, Wu Z, Liu S, Yao X. Current strategies for exosome cargo loading and targeting delivery. Cells. 2023;12:1416. doi:10.3390/cells12101416
65. Brancolini A, Vago R. Investigating the potential of extracellular vesicles as delivery systems for chemotherapeutics. Biomedicines. 2024;12:2863. doi:10.3390/biomedicines12122863
66. Guo Z, Zhang Y, Gong Y, et al. Antibody functionalized curcuma-derived extracellular vesicles loaded with doxorubicin overcome therapy-induced senescence and enhance chemotherapy. J Control Release. 2025;379:377–389. doi:10.1016/j.jconrel.2025.01.029
67. Lu Y, Zhang W, Zeng P, et al. Light modulates plant-derived extracellular vesicle properties: a photosensitive-responsive nanodelivery system. Discover Nano. 2025;20:86. doi:10.1186/s11671-025-04266-y
68. Man F, Meng C, Liu Y, et al. The study of ginger-derived extracellular vesicles as a natural nanoscale drug carrier and their intestinal absorption in rats. AAPS Pharm Sci Tech. 2021;22:206. doi:10.1208/s12249-021-02087-7
69. Kocholatá M, Malý J, Kříženecká S, Janoušková O. Diversity of extracellular vesicles derived from calli, cell culture and apoplastic fluid of tobacco. Sci Rep. 2024;14:30111. doi:10.1038/s41598-024-81940-8
70. Liu H, Liu J, Ji S, Peng S, Zhou L, Liu W. Effects of Orange variety on the physiochemical properties of self-secretory extracellular vesicle and its application potential as nutrient-rich beverage. Food Res Int. 2025;219:116972. doi:10.1016/j.foodres.2025.116972
71. Mammadova R, Maggio S, Fiume I, et al. Protein biocargo and anti-inflammatory effect of tomato fruit-derived nanovesicles separated by density gradient ultracentrifugation and loaded with curcumin. Pharmaceutics. 2023;15:333.
72. Cui L, Perini G, Palmieri V, De Spirito M, Papi M, Vasilyev OS. Plant-derived extracellular vesicles as a novel frontier in cancer therapeutics. Nanomaterials. 2024;15:14. doi:10.3390/nano15010014
73. Cui L, Perini G, Minopoli A, Palmieri V, De Spirito M, Papi M. Plant-derived extracellular vesicles as a natural drug delivery platform for glioblastoma therapy: a dual role in preserving endothelial integrity while modulating the tumor microenvironment. Int J Pharmaceut. 2025;10:100349. doi:10.1016/j.ijpx.2025.100349
74. Shen J, Wei T, Li M, et al. Aloe vera-derived extracellular vesicle-like particles suppress pancreatic carcinoma progression through triggering pyroptosis via ROS-GSDMD/E signaling pathway. Chin Med. 2025;20:101. doi:10.1186/s13020-025-01153-7
75. Kim K, Jung JH, Yoo HJ, et al. Anti-metastatic effects of plant sap-derived extracellular vesicles in a 3D microfluidic cancer metastasis model. J Funct Biomater. 2020;11:49. doi:10.3390/jfb11030049
76. Yeat NY, Chen RH. Extracellular vesicles: biogenesis mechanism and impacts on tumor immune microenvironment. J Biomed Sci. 2025;32:85. doi:10.1186/s12929-025-01182-2
77. Reale A, Khong T, Spencer A. Extracellular vesicles and their roles in the tumor immune microenvironment. J Clin Med. 2022;11:6892. doi:10.3390/jcm11236892
78. Yang J, Bahcecioglu G, Zorlutuna P. The extracellular matrix and vesicles modulate the breast tumor microenvironment. Bioengineering. 2020;7:7. doi:10.3390/bioengineering7010007
79. Qiang W, Li J, Ruan R, et al. Plant-derived extracellular vesicles as a promising anti-tumor approach: a comprehensive assessment of effectiveness, safety, and mechanisms. Phytomedicine. 2024;130:155750. doi:10.1016/j.phymed.2024.155750
80. Bellucci M, Pompa A, Lousa CDM, et al. Human Indoleamine 2,3-dioxygenase 1 (IDO1) expressed in plant cells induces kynurenine production. Int J Mol Sci. 2021;22:5102. doi:10.3390/ijms22105102
81. Song Y, Kong H, Oh S, Kim SB. Plant-derived extracellular vesicles as nanocarriers for combination therapy enhancing paclitaxel-based regimens in breast cancer. BMB Rep. 2025;58:53–63. doi:10.5483/BMBRep.2024-0193
82. Qin Y, Xu L, Guan YY, Liu H. Multifunctionalized protein-based drug delivery system for inhibition of tumor growth and progression. ACS Appl Bio Mater. 2020;3:3196–3202. doi:10.1021/acsabm.0c00190
83. Cao J, Pan Q, Bei S, et al. Concise nanoplatform of phycocyanin nanoparticle loaded with docetaxel for synergetic chemo-sonodynamic antitumor therapy. ACS Appl Bio Mater. 2021;4:7176–7185. doi:10.1021/acsabm.1c00745
84. Chen H, Qu H, Lu B, et al. A metal-coordination stabilized small-molecule nanomedicine with high drug-loading capacity and synergistic photochemotherapy for cancer treatment. Nanoscale. 2024;16:14734–14747. doi:10.1039/D4NR02101K
85. Guo C, Su Y, Cheng Z, et al. Novel ROS-responsive marine biomaterial fucoidan nanocarriers with AIE effect and chemodynamic therapy. Int J Biol Macromol. 2022;202:112–121. doi:10.1016/j.ijbiomac.2022.01.060
86. Yang Y, Wang P, Ji Z, Xu X, Zhang H, Wang Y. Polysaccharide‑platinum complexes for cancer theranostics. Carbohydrate Polymers. 2023;315:120997. doi:10.1016/j.carbpol.2023.120997
87. Wang X, Wu B, Sun G, et al. Selenium biofortification enhanced miR167a expression in broccoli extracellular vesicles inducing apoptosis in human pancreatic cancer cells by targeting IRS1. Int J Nanomed. 2023;18:2431–2446. doi:10.2147/IJN.S394133
88. López de Las Hazas MC, Tomé-Carneiro J, Pozo-Acebo LD, et al. Therapeutic potential of plant-derived extracellular vesicles as nanocarriers for exogenous miRNAs. Pharmacol Res. 2023;198:106999. doi:10.1016/j.phrs.2023.106999
89. Castelli G, Logozzi M, Mizzoni D, et al. Ex vivo anti-leukemic effect of exosome-like grapefruit-derived nanovesicles from organic farming-the potential role of ascorbic acid. Int J Mol Sci. 2023;24:15663. doi:10.3390/ijms242115663
90. Yin Q, Sun B, Qi X, et al. Anticancer activity of Momordica charantia-derived extracellular vesicle-like particle against cervical cancer. Tissue Cell. 2025;95:102897. doi:10.1016/j.tice.2025.102897
91. Wang F, Yuan M, Shao C, Ji N, Zhang H, Li C. Momordica charantia-derived extracellular vesicles provide antioxidant protection in ulcerative colitis. Molecules. 2023;28:6182.
92. Yang M, Luo Q, Chen X, Chen F. Bitter melon derived extracellular vesicles enhance the therapeutic effects and reduce the drug resistance of 5-fluorouracil on oral squamous cell carcinoma. J Nanobiotechnol. 2021;19:259. doi:10.1186/s12951-021-00995-1
93. Ye C, Yan C, Bian SJ, et al. Momordica charantia L.-derived exosome-like nanovesicles stabilize p62 expression to ameliorate doxorubicin cardiotoxicity. J Nanobiotechnol. 2024;22:464. doi:10.1186/s12951-024-02705-z
94. Kugeratski FG, McAndrews KM, Kalluri R. Multifunctional applications of engineered extracellular vesicles in the treatment of cancer. Endocrinology. 2021;162. doi:10.1210/endocr/bqaa250
95. Wang F, Li L, Deng J, et al. Lipidomic analysis of plant-derived extracellular vesicles for guidance of potential anti-cancer therapy. Bioact Mater. 2025;46:82–96. doi:10.1016/j.bioactmat.2024.12.001
96. Lai WY, Chuang CW, Huang YC, Huang CJ. Therapeutic potential of plant-derived small extracellular vesicles in sepsis: a network meta-analysis. Pharmacol Res. 2025;217:107795. doi:10.1016/j.phrs.2025.107795
97. Sileo L, Cavaleri MP, Lovatti L, Pezzotti G, Ferroni L, Zavan B. Dermatologically tested apple-derived extracellular vesicles: safety, anti-aging, and soothing benefits for skin health. J Cosmet Dermatol. 2025;24:e70254. doi:10.1111/jocd.70254
98. Zhao M, Chen X, Wang W, et al. Sea buckthorn-derived extracellular vesicles foster bone regeneration through aau-miR168-mediated pathways. Stem Cell Res Ther. 2025;16:281. doi:10.1186/s13287-025-04373-8
99. Ekanayake G, Piibor J, Midekessa G, et al. Systematic characterization of extracellular vesicles from potato (Solanum tuberosum cv. Laura) roots and peels: biophysical properties and proteomic profiling. Front Plant Sci. 2024;15:1477614. doi:10.3389/fpls.2024.1477614
100. Ipinmoroti AO, Turner J, Bellenger EJ, et al. Characterization of cannabis strain-plant-derived extracellular vesicles as potential biomarkers. Protoplasma. 2023;260:1603–1606. doi:10.1007/s00709-023-01870-6
101. Wu P, Wu W, Zhang S, et al. Therapeutic potential and pharmacological significance of extracellular vesicles derived from traditional medicinal plants. Front Pharmacol. 2023;14:1272241. doi:10.3389/fphar.2023.1272241
102. Lou K, Luo H, Jiang X, Feng S. Applications of emerging extracellular vesicles technologies in the treatment of inflammatory diseases. Front Immunol. 2024;15:1364401. doi:10.3389/fimmu.2024.1364401
103. Karamanidou T, Tsouknidas A. Plant-derived extracellular vesicles as therapeutic nanocarriers. Int J Mol Sci. 2021;23:191. doi:10.3390/ijms23010191
104. Lian MQ, Chng WH, Liang J, et al. Plant-derived extracellular vesicles: recent advancements and current challenges on their use for biomedical applications. J Extracell Vesicles. 2022;11:e12283. doi:10.1002/jev2.12283
105. Kalishwaralal K, Nazeer AA, Induja DK, Keerthana CK, Shifana SC, Anto RJ. Enhanced extracellular vesicles mediated uttroside B (Utt-B) delivery to Hepatocellular carcinoma cell: pharmacokinetics based on PBPK modelling. Biochem Biophys Res Commun. 2024;703:149648. doi:10.1016/j.bbrc.2024.149648
106. Garaeva L, Kamyshinsky R, Kil Y, et al. Delivery of functional exogenous proteins by plant-derived vesicles to human cells in vitro. Sci Rep. 2021;11:6489. doi:10.1038/s41598-021-85833-y
107. Rosenkrans ZT, Thickens AS, Kink JA, et al. Investigating the in vivo biodistribution of extracellular vesicles isolated from various human cell sources using positron emission tomography. Mol Pharm. 2024;21:4324–4335. doi:10.1021/acs.molpharmaceut.4c00298
108. Zhang X, Long Z, Qin Z, et al. A detailed evaluation of the advantages among extracellular vesicles from three cell origins for targeting delivery of celastrol and treatment of glioblastoma. Int J Pharm. 2024;667:125005. doi:10.1016/j.ijpharm.2024.125005
109. Sah NK, Arora S, Sahu RC, Kumar D, Agrawal AK. Plant-based exosome-like extracellular vesicles as encapsulation vehicles for enhanced bioavailability and breast cancer therapy: recent advances and challenges. Med Oncol. 2025;42:184. doi:10.1007/s12032-025-02720-6
110. Zhu Y, Zhao J, Ding H, et al. Applications of plant-derived extracellular vesicles in medicine. MedComm. 2024;5:e741. doi:10.1002/mco2.741
111. Liu H, Liu Y, Peng S, et al. Colonic delivery and controlled release of curcumin encapsulated within plant-based extracellular vesicles loaded into hydrogel beads. Food Res Int. 2025;202:115540. doi:10.1016/j.foodres.2024.115540
112. Muhammad Z, Muhammad SA, Abbas AY, et al. Isolation and characterization of medicinal plant-based extracellular vesicles as nano delivery systems for ascorbic acid. J Microencapsul. 2025;42:120–131. doi:10.1080/02652048.2024.2443430
113. Bhom N, Somandi K, Ramburrun P, Choonara YE. Extracellular nanovesicles as neurotherapeutics for central nervous system disorders. Expert Opin Drug Delivery. 2025;22:69–84. doi:10.1080/17425247.2024.2440099
114. Kang J, Wen J, Chen H, Zhu C, Bai Y. Progress of research on engineered extracellular vesicles from different sources for disease treatment. Histol Histopathol. 2025;40:1501–1518. doi:10.14670/HH-18-903
115. Silva RM, Azevedo AM, Bonifácio VDB, Fernandez-Becerra C, Pinto SN. Unlocking the potential of extracellular vesicles: one stimulus away from clinical implementation. Biomater Sci. 2025;13:6483–6509.
116. Wei C, Zhang M, Cheng J, Tian J, Yang G, Jin Y. Plant-derived exosome-like nanoparticles – from Laboratory to factory, a landscape of application, challenges and prospects. Crit Rev Food Sci Nutr. 2025;65:4510–4528. doi:10.1080/10408398.2024.2388888
117. Rawat S, Arora S, Dhondale MR, Khadilkar M, Kumar S, Agrawal AK. Stability dynamics of plant-based extracellular vesicles drug delivery. J Xenobiot. 2025;15:55.
118. Boccia E, Alfieri M, Belvedere R, et al. Plant hairy roots for the production of extracellular vesicles with antitumor bioactivity. Commun Biol. 2022;5:848. doi:10.1038/s42003-022-03781-3
119. Tong Q, Li K, Huang F, et al. Extracellular vesicles hybrid plasmid-loaded lipid nanovesicles for synergistic cancer immunotherapy. Mater Today Bio. 2023;23:100845. doi:10.1016/j.mtbio.2023.100845
120. Fan Z, Wang Z, Zhang H, et al. Extracellular vesicles derived from mature dendritic cells loaded with cDC1-specific chemokine XCL1 combined with chemotherapy-induced ICD for the treatment of castration-resistant prostate cancer. Cancer Immunol Immunother. 2025;74:242. doi:10.1007/s00262-025-04070-8